Le Réseau Mammite rassemble plusieurs chercheurs canadiens et internationaux
engagés à diminuer l’incidence de la mammite, à réduire les pertes économiques et à maintenir la qualité du lait par une recherche concertée et un transfert efficace et rapide des résultats aux utilisateurs.
Le Réseau
Notre Mission

Notre empreinte
sur les fermes du Canada

01
Soutien à la durabilité des fermes
en limitant les pertes économiques causées par la mammite

02
Réduction de l’usage des antibiotiques

03
Lutte à
l’antibio-résistance
et protection de la santé publique

04
Amélioration du bien-être des vaches laitières
et diminution de la souffrance animale
L’envergure du Réseau:
universités canadiennes
centre fédéral de recherche
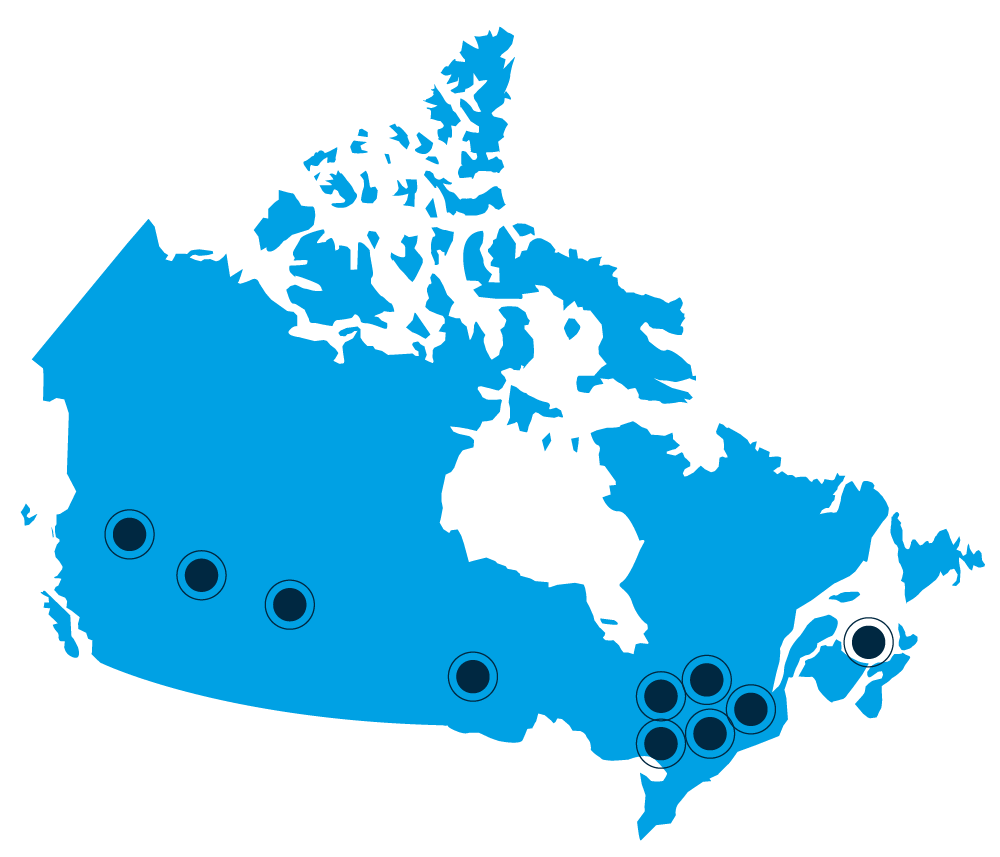
Colombie-Britannique
• University of British Columbia
Alberta
• University of Calgary
Saskatchewan
• University of Saskatchewan et VIDO
Ontario
• University of Guelph
Québec
• Université Laval
• Université McGill
• Université de Montréal
• Université de Sherbrooke
• Centre de recherche et de développement sur le bovin laitier et le porc – Sherbrooke
Île-du-Prince-Édouard
• University of Prince Edward Island
Structure du Réseau
Les activités de recherche du RM sont encadrées et évaluées par un Comité scientifique. Les membres font les recommandations aux chercheurs pour assurer un suivi adéquat des projets de recherche.
Ce comité est composé des directeurs, des responsables de thèmes, de représentants de l’industrie et d’experts internationaux.
Une équipe administrative, située à la Faculté de médecine vétérinaire de l’Université de Montréal à Saint-Hyacinthe au Québec, s’occupe de l’administration des fonds de recherche, du suivi des projets et du développement des activités de transfert des connaissances.
Organigramme du Réseau
Directeur
Simon Dufour
Gestionnaire
Josée Labrie
Agente de transfert
Ibtissem Doghri
Responsables des axes
Thème pathogènes
G. Keefe, U.PEI, Calgary
Thème animal
P. Lacasse, AAC
Thème environnement
H. Barkema, U. Calgary
Comités
Comité exécutif
Directeur, responsables des axes et gestionnaire du réseau
Comité scientifique
Directeur, responsables des axes et gestionnaire du réseau, chercheurs du réseau et producteur
Connaissez-vous vraiment tout sur la mammite bovine?
MOOC 1
MOOC 2
MOOC 3
Nos partenaires
Nos projets de recherche sont soutenus par une contribution de l’initiative de la Grappe de recherche laitière (Les Producteurs laitiers du Canada, Agriculture et Agroalimentaire Canada, le Réseau laitier canadien et la Commission canadienne du lait).
